Principios del Ligamiento
Publicado por: Naileth Castro
El principio de Mendel según el cual los genes que controlan diferentes caracteres son heredados de forma independiente uno de otro es cierto sólo cuando los genes existen en cromosomas diferentes. El genetista estadounidense Thomas Hunt Morgan y sus colaboradores demostraron en una serie amplia de experimentos con moscas de la fruta (que se reproducen con gran velocidad), que los genes se disponen de forma lineal en los cromosomas y que cuando éstos se encuentran en el mismo cromosoma, se heredan como una unidad aislada mientras que el cromosoma permanezca intacto. A los genes heredados de esta manera se dice que “están ligados”.
Sin embargo, Morgan y su grupo observaron también que este ligamiento rara vez es completo. Las combinaciones de los alelos de cada progenitor pueden reorganizarse. Durante la meiosis, una pareja de cromosomas homólogos puede intercambiar material durante lo que se denomina “recombinación o entrecruzamiento”. El entrecruzamiento se produce generalmente al azar a lo largo de los cromosomas, de modo que la frecuencia de recombinación entre dos genes depende de la distancia que los separe en el cromosoma. En consecuencia los científicos pueden trazar o dibujar mediante experimentos de reproducción apropiados, las posiciones relativas de los genes a lo largo del cromosoma o mapa génico.
Basándose en la Teoría Cromosómica de la Herencia enunciada por Sutton y cuyos aspectos esenciales son:
- Los genes están ubicados en los cromosomas
- La ordenación de los mismos es lineal
- Al fenómeno genético de la recombinación le corresponde fenómeno citológico de intercambio de segmentos cromosómicos.
Morgan propuso que Existen parejas génicas situadas sobre el mismo par de cromosomas homólogos, llamando a este fenómeno ligamiento.

Referencia:
http://www.vet.unicen.edu.ar/…/4-Ligamiento%20y%20Recombinacion.pdf
Ligamiento y Recombinación
Publicado por: Naileth Castro
Los genes que se encuentran en distintos cromosomas se distribuyen en los gametos independientemente uno del otro. Sin embargo, los genes que se encuentran en el mismo cromosoma tienden a permanecer juntos; es decir, a no sufrir separaciones ni combinaciones al azar durante la formación de los gametos.
Al analizar los resultados de un cruce de prueba a individuos dihíbridos se observarán resultados diferentes, dependiendo de la ubicación de los genes, es decir: en el mismo o en distintos cromosomas.
Genes con distribución independiente
- Los genes que se encuentran en distintos cromosomas se distribuyen independientemente, por lo tanto del apareamiento AaBb x aabb se obtendrá una proporción esperada en el cruce de prueba de 1:1:1:1
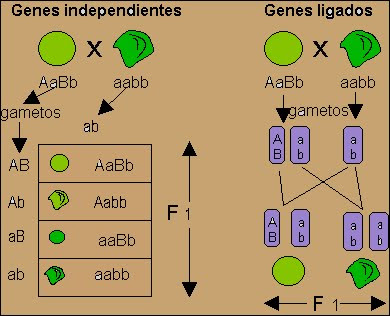
Genes completamente ligados o Ligamiento Completo
Los productos de la meiosis (gametos AB y ab) presentan los genes ligados o unidos de la misma forma que se encuentran en el progenitor sometido a prueba. Son el resultado de no haber sufrido entrecruzamiento y se denominan “gametos tipo progenitor o parental”.
Genes parcialmente ligados o Ligamiento Incompleto y Recombinación
- Los genes ligados no siempre permanecen juntos, debido a que las cromátides no hermanas de cromosomas homólogos pueden intercambiar entre ellos segmentos de longitud variable durante la profase meiótica produciendo “gametos de tipo recombinantes” por virtud del entrecruzamiento. Cabe aclarar que en las meiosis que ocurre entrecruzamiento y recombinación además de los gametos de tipo progenitor se obtienen los gametos tipo recombinantes que constituyen nuevas combinaciones de los genes ligados en los progenitores.

Meiosis y Recombinación
Durante la meiosis cada cromosoma se duplica formando dos cromátides hermanas idénticas, el par de cromosomas homólogos se aparea (sinapsis) y se produce el entrecruzamiento entre cromátides no hermanas. Este último proceso requiere del rompimiento y la reunión de sólo dos de las cuatro bandas en cualquier punto del cromosoma.
Los productos de la meiosis (gametos AB y ab), presentan los genes ligados de la misma forma que se encuentran en los progenitores, resultan de las cromátides que no sufrieron entrecruzamiento y se denominan “gametos tipo progenitor o parental”.
Los otros dos productos meiótico (gametos Ab y aB), resultantes del entrecruzamiento, constituyen nuevas combinaciones de los genes ligados originalmente en los progenitores y se denominan “gametos tipo recombinantes”.


Fase de Acoplamiento
En los dihíbridos o dobles heterocigotos, para dos loci ligados, puede ocurrir que los dos alelos dominantes o tipo común estén en un cromosoma y los dos recesivos o mutantes en el otro. En este caso se dice que las relaciones de enlace son de acoplamiento o “cis”.
Fase de Repulsión
Cuando el alelo dominante de un locus y el recesivo del otro ocupan el mismo cromosoma la relación es de repulsión o “trans”. Los gametos recombinantes y progenitores serán diferentes en cada caso.
La recombinación, definida en relación a la meiosis, es el proceso que genera un producto haploide cuyo genotipo difiere de los dos genotipos progenitores (haploides) que formaron la célula (2n) que inició la meiosis. El producto así generado se denomina recombinante.

Hay dos tipos de recombinación, las que producen recombinantes por método absolutamente diferentes:
- Intercromosómica
- Intracromosómica
Recombinación intercromosómica.
Es la que se produce mediante la distribución independiente de Mendel. Las dos clases recombinantes o nuevos fenotipos constituyen el 50% de los descendientes, 25% de cada tipo. Si encontramos esta frecuencia podemos inferir que las parejas génicas segregan independientemente.
Recombinación intracromosómica.
Se produce por entrecruzamiento. Esto ocurre entre cualquiera de dos cromátides no hermanas. Esto no sucede en todas las meiosis, pero cuando lo hace, la mitad de esos productos son recombinantes y la otra mitad serán gametos tipo progenitor. Las meiosis sin entrecruzamiento entre dos loci producirán sólo genotipos parentales para esas parejas génicas.
La recombinación intracromosómica se pone de manifiesto en la aparición de una frecuencia de recombinación menor al 50%. El ligamiento físico entre las combinaciones génicas parentales impide la libre distribución de los genes (distribución independiente), la que genera una frecuencia de recombinación del 50%.

Para mayor información, visualiza el siguiente video:
Referencia:
http://www.vet.unicen.edu.ar/…/4-Ligamiento%20y%20Recombinacion.pdf
Coincidencia e Interferencia
Publicado por: Jesús Camacaro
En genética la Interferencia se expresa como la medida del grado de los entrecruzamientos, que se calcula sustrayendo el coeficiente de coincidencia del valor 1. Mientras que el coeficiente de coincidencia se define en términos generales; como el cociente entre el número de dobles entrecruzamientos observados y el esperado.
Se sabe que en la mayoría de los organismos superiores, la formación de un quiasma (Ver Imagen N° 1), reduce la probabilidad de formación de otro quiasma en la región inmediatamente adyacente del cromosoma. Puede considerarse que esta reducción en la formación de entrecruzamientos entre cromáticas no hermanas, se deba a la incapacidad física de las mismas, para plegarse sobre sí dentro de cierta distancia mínima.

Imagen N° 1. Formación de Quiasma.
Fuente:http://biologiabi.blogspot.com/2011/07/32-cruces-dihibridos-y-ligamiento-de.html
Si el entrecruzamiento de una pareja de cromátidas no afectase el entrecruzamiento de una pareja vecina, se espera que los dobles entrecruzamientos se presenten con una frecuencia igual al producto de ambas frecuencias. Sin embargo si la frecuencia observada es menor a la esperada, aparentemente el entrecruzamiento en una región interfiere con el entrecruzamiento en una región vecina.
El resultado neto de esta interferencia es la observación de que se presentan menos entrecruzamientos dobles, de los que habría de esperarse de acuerdo con las distancias de mapa establecidas. La fuerza de la interferencia, varía en diferentes segmentos del cromosoma y se expresa comúnmente en términos del coeficiente de coincidencia, o el cociente entre los entrecruzamientos dobles observados y los esperados.
La Fórmula del Coeficiente de Coincidencia se expresa de la siguiente manera:

La coincidencia es el complemento de la interferencia (Coincidencia + Interferencia = 1.0), en función a esto cuando la interferencia es completa (1.0), no se observarán entrecruzamientos dobles y la coincidencia tendrá un valor de cero. Por el contrario cuando se observan todos los entrecruzamientos dobles esperados, la coincidencia es uno y la interferencia cero.
Ejemplo:
Dadas las distancias de mapa A – B – 20 u.m y B – C de 30 u.m, entonces se espera 0.2X0.3 = 0.06 o 6% de entrecruzamientos dobles si no hay interferencia. Suponga que se observa 1.3% de entrecruzamientos dobles en una cruza de prueba experimental.
Coincidencia = 1.3/6.0 = 0.2.
Esto significa simplemente que sólo se observa el 20% de los entrecruzamientos dobles que se esperan sobre la base de combinar las probabilidades independientes (distancias de mapa).
Interferencia = 1.0 – 0.2 = 0.8.
Así, el 80% de los entrecruzamientos dobles no se formó debido a la interferencia.
Referencias
Griffith, A Miller, J., Suzuki, D. Lewontin, R. y Gealbart, W. (1998). Introducción al Análisis Genético. España: Internacional,Mc Garw Hill.
Rojas, G. (2009). La genética mendeliana. La Teoría cromosómica de la Herencia. Las Mutaciones. . [Documento en línea] Disponible: http://www.preparadores.eu/temamuestra/Secundaria/ByG.pdf. [Consulta: 2015, Enero 24].
Entrecruzamiento de Dos Puntos
Publicado por: Angélica Bastidas
El entrecruzamiento cromosómico se refiere a la recombinación entre los cromosomas apareados heredado de uno de los padres, generalmente ocurre durante la meiosis. Durante la profase I, las cuatro cromátides disponibles están estrechamente posicionadas una con respecto a la otra. Mientras en esta formación, los sitios homólogos en las dos cromátides pueden coincidir entre sí, y pueden intercambiar información genética.

Como la recombinación puede producirse con baja probabilidad en cualquier lugar del cromosoma, la frecuencia de recombinación entre dos puntos depende de su distancia. Por lo tanto, para genes suficientemente distantes en el mismo cromosoma la cantidad de recombinación es lo suficientemente alta para destruir la correlación entre alelos.
Se denomina así a la recombinación entre los cromosomas apareados, generalmente durante la meiosis. Durante la profase I, en la sub-fase de paquitene, las cuatro cromáticas disponibles están estrechamente posicionadas una con respecto a la otra. En esta disposición los sitios homólogos en las dos cromáticas pueden coincidir entre sí, y pueden intercambiar información genética.
Como la recombinación puede producirse en cualquier lugar del cromosoma, la frecuencia de recombinación entre dos puntos depende de la distancia entre ambos. Por lo tanto, para genes suficientemente distantes en el mismo cromosoma la frecuencia de recombinación es lo suficientemente alta para destruir la correlación entre alelos recombinantes.
Entrecruzamiento cromosómico
El Entrecruzamiento cromosómico. Es el proceso por el cual dos cromosomas se aparean e intercambian secciones de su ADN Durante la meiosis. Tiene como resultado una combinación nueva de material genético en el gameto La sinapsis comienza antes de que se desarrolle el complejo sinaptonémico, y no está completo hasta cerca del final de la profase 1. El entrecruzamiento usualmente se produce cuando se aparean las regiones en las rupturas del cromosoma y luego se reconectan al otro cromosoma. El resultado de este proceso es un intercambio de genes, llamado recombinación genética. Los entrecruzamientos cromosómicos también suceden en organismos asexuales y en células somáticas, ya que son importantes formas de reparación del ADN.
Para mayor información, visualiza el siguiente video:
Entrecruzamiento de Tres Puntos
Publicado por: Jesús Camacaro
Cuando la distancia física entre dos Loci es lo suficientemente grande, pueden ocurrir dos (o incluso más) entrecruzamientos en la región que los separa. Para la identificación de los dobles entrecruzamientos es preciso seguir la segregación de un tercer locus en el estudio de recombinación. Estos cruces de tres puntos incluyen un par alélico localizado en el espacio interior de los marcadores genéticos, de forma tal que los intercambios dobles en la región podrán ser detectados, cuando los entrecruzamientos flanqueen a este gen. La identificación de los tipos doble recombinantes, convierte a los cruces de tres puntos no solo en un método eficiente para obtener datos de recombinación, sino para determinar el orden de los genes de un modo simple.
Al analizar lo que ocurre cuando se da un entrecruzamiento entre tres genes que están ligados (Ver Imagen N° 1), se debe tener en cuenta que los genes muestran una configuración de acoplamiento (alelos dominantes en un cromosoma A-B-C y recesivos en otros a-b-c), pudiéndose producir tres tipos de entrecruzamiento; dos entrecruzamientos simpes y un entrecruzamiento doble.

Imagen N° 1. Entrecruzamiento de Tres Puntos.
En los cromosomas recombinantes que resultan del entrecruzamiento doble, los alelos de los extremos son los mismos que en los cromosomas no recombinantes, pero el alelo del medio es diferente. Este resultado revela un dato importante respecto al orden de los genes. En la progenie que resulta de un entrecruzamiento doble, el alelo del medio es el único que debería diferir de los alelos presentes en la progenie no recombinante.
Como regla general, cuando participan tres pares de genes se deberían producir ocho recombinaciones. Si ocurre un solo entrecruzamiento, se origina un simple recombinante; si hay dos entrecruzamientos, se denomina un doble entrecruzamiento y de esta forma se separa al gen que se encuentra en el medio.
Se ha observado que la frecuencia de dobles entrecruzamientos es baja, porque resulta muy difícil separar al gen que se encuentra en el medio de los otros dos genes. Por esta razón, los dobles entrecruzamientos se producen en menor proporción que la esperada.
Pasos Claves para hacer un Mapeo de Genes de un Cruzamiento de Tres Puntos
- Anotar los fenotipos de los números de la progenie producida en el cruzamiento de tres puntos. Utilizar símbolos que representen a los alelos para las características.
- Anotar los genotipos de los progenitores originales, a través de los cuales se produjo el individuo triplemente heterocigoto. Esto del cruzamiento de prueba y si también se conoce, anotar también la disposición de los alelos (acoplamiento y repulsión), en los cromosomas del individuo.
- Determinar que clase fenotípica entre la progenie son no recombinantes y cuáles son las que resultaron de entrecruzamientos dobles. Las no recombinantes serán las dos clases fenotípicas más frecuentes y las de entrecruzamientos dobles serán las dos clases menos frecuentes.
- Determinar cuál es el locus que se encuentra en el medio. Comprar los alelos presentes en los entrecruzamientos dobles con aquellos presentes en los no recombinantes; cada clase de entrecruzamientos dobles debe coincidir con una de las clases no recombinantes respecto de dos locis y diferir respecto a un locus. Este último será el que se encuentra en el medio.
- Volver a anotar los fenotipos con el orden correcto.
- Determinar en qué ubicaciones pueden haber ocurrido los entrecruzamientos para dar origen a los fenotipos de la progenie. Compara cada fenotipo con el fenotipo de la progenie no recombinante.
- Determinar las frecuencias de recombinación. Sumar las cifras de la progenie que posee un cromosoma con un entrecruzamiento entre un par de loci. Sumar los entrecruzamientos dobles a esta cifra. Dividir el resultado por el número total de la progenie del cruzamiento y multiplicarlo por 100%, el resultado obtenido corresponde a la frecuencia de recombinación entre los loci, que es igual a la distancia de mapa.
- Construir un mapa de los tres Loci. Indicar cuál es el que se encuentra en el medio y marcar la distancia entre ellos.
- Determinar el coeficiente de interferencia y de coincidencia. Utilizando la ecuación respectiva.
Referencias
Pierce, B. (2009). Genética Un Enfoque Conceptual. España: Editorial Medica Panamericana, S.A.
Griffith, A Miller, J., Suzuki, D. Lewontin, R. y Gealbart, W. (1998). Introducción al Análisis Genético. España: Internacional,Mc Garw Hill.
Cromosomas, Cariotipo y Mutaciones. Relación entre sí
Publicado por: Evacaroly Colmenárez
Los cromosomas son estructuras que se encuentran en el centro (núcleo) de las células que transportan fragmentos largos de ADN. Cabe destacar que el ADN es el material que contiene los genes y es el pilar fundamental del cuerpo humano.
En este mismo sentido los cromosomas se organizan en pares. Normalmente en las células humanas los cromosomas forman 23 pares de cromosomas, en donde 22 son autosómicos y el par 23 se conocen como cromosomas sexuales, y es el que determina el sexo, es decir si usted es niño o niña. Así pues que la mujer consta de un par cromosómico XX, mientras que el hombre un cromosoma X y otro Y (XY). En otras palabras La madre le aporta un cromosoma X al hijo, mientras que el padre puede contribuir ya sea con un cromosoma X o con un cromosoma Y. Es el cromosoma del padre el que determina si el bebé es un niño o una niña.
En la siguiente imagen se puede observar las partes de un cromosoma

Por otra parte el Cariotipo es el ordenamiento de los cromosomas matafásicos, de acuerdo con su tamaño y morfología (metacéntricos, submetacéntricos, telocéntricos, subtelocéntricos y acrocéntricos). Cabe destacar que el cariotipo describe las características de sus cromosomas mediante un código
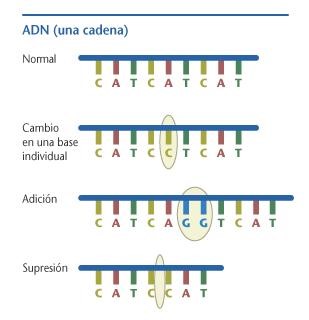
Una Mutación del Latín mutare=cambiar, es un cambio en la información contenida en el ADN de las células. Para que sea heredable tiene que ocurrir en las células sexuales: óvulos y espermatozoides. Es importante señalar que en la naturaleza las mutaciones se producen al azar, pero pueden ser estimuladas mediante agentes mutagénicos, como las radiaciones y sustancias químicas. La existencia de varios alelos para un mismo gen se debe a la existencia de mutaciones.
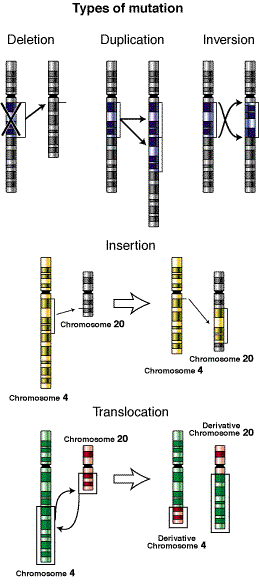
Cabe destacar que cuando una mutación afecta solo a un gen se le denomina mutación genética; mientras que cuando la mutación afecta la estructura de uno o varios cromosomas se le denomina mutación cromosómica; y cuando una o varias mutaciones provoca alteraciones en todo el material genético se le denomina mutación genómica
Podemos decir que las mutaciones son el origen de la diversidad genética
En la siguiente imagen se puede observar algunos tipos de mutaciones
Mutación del ADN
Mutaciones cromosómicas
Trisonomía del par 21 de cromosomas

Mutación genética (Anemia Falciforme)

A continuación se muestran unos videos para ampliar la información.
Referencias
- Adolph, K. (ed.) (1988)Chromosomes and chromatin 1—3 Boca Raton FL: CRC Press.
- Stewart, A. (1990)The functional organization of chromosomes and the nucleus, a special issue. Trends Genet. (6):377—379
Unidades de Mapa y Frecuencia de Recombinación
Publicado por: Antony Vásquez
Thomas Hunt Morgan
(25 de septiembre 1866 — 4 de diciembre 1945) fue un genetista estadounidense. Estudió la historia natural, zoología, y macromutación en la mosca de la fruta Drosophila melanogaster.
Sus contribuciones científicas más importantes fueron en el campo de la Genética. Fue galardonado con el Premio Nobel de Fisiología y Medicina en 1933 por la demostración de que los cromosomas son portadores de los genes, lo que se conoce como la teoría cromosómica de Sutton y Boveri. Gracias a su trabajo, Drosophila melanogaster se convirtió en uno de los principales organismos modelo en Genética.

Morgan nació en Lexington, Kentucky, hijo de Charlton Hunt Morgan y Ellen Key Howard, y sobrino del general confederado John Hunt Morgan.Tuvo una juventud dura según Carl Phomson en su libro la vida de la Biología 4.
Morgan se graduó en la Universidad de Kentucky en 1886. Recibió el doctorado de la Universidad Johns Hopkins en 1890. Siguiendo los pasos de William E. Castle, comenzó a trabajar en el desarrollo embrionario de Drosophila melanogaster (la mosca de la fruta) en la Universidad de Columbia, donde se interesó por el problema de la herencia. Las teorías de Gregor Mendel acababan de ser redescubiertas en 1900 y Morgan estaba interesado en estudiar su aplicación a los animales.
En 1910, descubrió un mutante de ojos blancos entre individuos de estirpe silvestre de ojos rojos. La progenie del cruzamiento de un macho de ojos blancos con una hembra de ojos rojos presentó ojos rojos, lo que indicaba que el carácter «ojos blancos» era recesivo. Morgan denominó white al gen correspondiente, iniciando así la tradición de nombrar a los genes según el fenotipo causado por sus alelos mutantes. Al cruzar estas moscas entre sí, Morgan se percató de que sólo los machos mostraban el carácter «ojos blancos». De sus experimentos, concluyó que algunos caracteres se heredan ligados al sexo, que el gen responsable del carácter residía en el cromosoma X, y que probablemente otros genes también residían en cromosomas específicos. Él y sus estudiantes contaron las características de miles de moscas y estudiaron su herencia. Empleando la recombinación de los cromosomas, Morgan y Alfred Sturtevant prepararon un mapa con la localización de los genes en el cromosoma. Morgan y sus estudiantes también escribieron el libro Mechanisms of Mendelian Heredity. Morgan se trasladó a CalTech en 1928. Morgan murió en Pasadena, California.
Obra
Ilustración de Thomas Hunt Morgan de la recombinación genética (1916).
En Mechanisms of Mendelian Heredity Morgan se opuso radicalmente a la herencia citoplasmática, estableciendo la recombinación genética como el mecanismo evolutivo fundamental.
Legado
El legado de Morgan a la Genética es muy importante. Algunos de los estudiantes de Morgan en Columbia y CalTech ganaron sus propios Premios Nobel, como George Wells Beadle, Edward B. Lewis y Hermann Joseph Muller. En honor de Morgan, la Genetics Society of America concede anualmente la Medalla «Thomas Hunt Morgan» para premiar las contribuciones significativas a la ciencia de la Genética. Eric Kandel, galardonado con el Premio Nobel, ha escrito sobre Morgan: «Así como las ideas de Darwin sobre la evolución de las especies dieron coherencia a la Biología del siglo XIX como ciencia descriptiva, los hallazgos de Morgan sobre los genes y su localización en los cromosomas ayudaron a convertir la Biología en una ciencia experimental.» La unidad de frecuencia de recombinación usada en cartografía genética, se denomina centiMorgan en su honor.
En el laboratorio de Morgan en la Universidad de Columbia realizó una larga y fructífera estancia José Fernández Nonídez. Nonídez fue uno de los pioneros españoles de la Genética, y difundió la teoría cromosómica de la herencia en España.
![Drosophila_melanogaster[1]](https://upelbiogenetica.wordpress.com/wp-content/uploads/2015/01/drosophila_melanogaster1.jpg)
Trabajos
Se reconoció la presencia de los cromosomas sexuales y de lo que se conoce en genética como “herencia ligada al sexo”. Demostró que los factores mendelianos (los genes) se disponían de forma lineal sobre los cromosomas. Los experimentos realizados por Morgan y colaboradores revelaron también la base genética de la determinación del sexo. Morgan continuó sus experimentos y demostró en su «Teoría de los genes» que los genes se encuentran unidos en diferentes grupos de encadenamiento, y que los alelos (pares de genes que afectan al mismo carácter) se intercambian o entrecruzan dentro del mismo grupo.
Experimentos
Thomas Hunt Morgan trabajó intensamente en un programa de reproducción y cruce de miles de moscas de la fruta en la Universidad de Nueva York en un cuarto que pasó a llamarse el Cuarto de la Moscas. Intentó hacer mutar las moscas con diversos medios (rayos X, centrifugadoras, etc).
La mosca de la fruta la cual posee 4 pares de cromosomas. Uno de esos pares se identificó como conteniendo cromosomas sexuales X y Y. Aplicó los principios mendelianos en las moscas. El estudio de herencia realizado por Morgan demostró la herencia ligada al sexo, y es una de las primeras evidencias que confirman la teoría cromosómica de la herencia basada en el cruzamiento.
En 1909, Morgan observó una mosca de la fruta (Drosophila melanogaster) con una mutación extraña a la que llamo “ojos blancos”, debido precisamente a la coloración de sus ojos (contraria a la normal, que es roja). Analizando esta mosca al microscopio Morgan descubrió que era un macho, y decidió usarlo como semental para así poder observar cómo iría pasando de generación en generación la nueva característica de ojos blancos.
Toda la descendencia de esta cruza resultó tener los ojos rojos, lo cual hizo sospechar a Morgan que algo raro había ocurrido, pues el color de los ojos del padre no podía haber desaparecido. Decidió entonces tomas a un par de “hijas moscas” y cruzarlas entre sí, simplemente para ver que pasaba. La sorpresa de Morgan fue muy grande, al observar que entre las moscas “nietas” había machos con los ojos blancos.

El problema entonces fue explicar qué había ocurrido durante la transmisión hereditaria para que el color de los ojos blancos sólo lo poseyeran los machos.
Morgan propuso la herencia ligada al sexo, es decir, la existencia de caracteres ligados al cromosoma sexual X de las hembras.
Posteriormente, Morgan encontró otras características que se heredaban de la misma manera, haciendo cada vez más sólida su idea de que estaban ligadas al cromosoma sexual. En ese momento Morgan adopta la palabra gen o genes para describir estos factores que se heredaban junto con el cromosoma X, argumentando que quizá estos genes estaban alineados formando parte de los cromosomas, los cuales, en su conjunto, formaban el acervo genético de los individuos y de las especies.
Conforme avanzaron sus investigaciones, encontraron más genes que estaban asociados con el cromosoma sexual, y más aún, encontraron factores que estaban localizados en los cromosomas I, II y III. Esto implicó necesariamente pensar que había una relación entre la transmisión de los cromosomas y la aparición de ciertos caracteres. Los genes eran transmitidos al mismo tiempo que el cromosoma, de tal forma que ciertos factores contenidos en los cromosomas darían un patrón de herencia en el que los genes que se encontraban en cada uno de ellos funcionaban como un grupo de ligamiento.
Así, se asociaron por primera vez los cromosomas con los genes y se determinó que estos últimos se comportaban de acuerdo con el comportamiento de los cromosomas durante la meiosis. Esto es lo que se conoce como la teoría cromosómica de la herencia.
La teoría cromosómica de la herencia dice que “los genes estaban en los cromosomas, y que, por lo tanto, los genes que se encontraban en el mismo cromosoma tienden a heredarse juntos, proponiendo para ellos el término «genes ligados». Según Morgan, los genes están en los cromosomas, su disposición es lineal, uno detrás de otro, y mediante el entrecruzamiento de las cromátidas homólogas se produce la recombinación genética.

Definiciones
Genética (del griego antiguo γενετικός /guennetikós/, ‘genetivo’, y este de γένεσις /guénesis/, ‘origen’) es el campo de la biología que busca comprender la herencia biológica que se transmite de generación en generación.
El estudio de la genética permite comprender qué es lo que exactamente ocurre en el ciclo celular, (replicar nuestras células y reproducción, (meiosis) de los seres vivos y cómo puede ser que, por ejemplo, entre seres humanos se transmiten características biológicas genotipo (contenido del genoma específico de un individuo en forma de ADN), características físicas fenotipo, de apariencia y hasta de personalidad.
Cartografía genética es una disciplina de la genética que, mediante varias técnicas, busca asignar a los distintos genes de un genoma su lugar físico en aquél. Existen dos variantes fundamentales de mapas: los genéticos, definidos mediante unidades de frecuencia de recombinación, y los físicos en los que las distancias entre loci se expresan en unidades de distancia en nucleótidos
Frecuencia de recombinación es un parámetro genético de cartografía genética, que, para dos loci dados, se emplea como indicador cuantitativo de la distancia que existe entre ambos. Su unidad, la unidad de mapa genético o centiMorgan(cM), en honor de Thomas Hunt Morgan, como la frecuencia de recombinación de 0,01, esto es, del 1%, lo que quiere decir que uno de cada 100 productos de meiosis es recombinante.
Centimorgan (abreviado cM) es la unidad de los mapas genéticos de ligamiento realizados por recombinación en eucariotas diploides. Se trata de una unidad indivisible: el prefijo centi-, en este caso, no significa centésima parte de Morgan. Equivale a un 1% de frecuencia de recombinación, o lo que es lo mismo, a 1 unidad de mapa.
Su nombre rinde honor a Thomas Hunt Morgan, genético estadounidense que estableció la teoría cromosómica de la herencia, descubrió la determinación cromosómica del sexo y realizó el primer mapa genético. Fue acuñado por Alfred Sturtevant, colaborador de Morgan, que empleó el prefijo centi- para darle un toque más «científico». Los investigadores estadounidenses no empleaban el sistema internacional por entonces.
Para mayor información, visualiza los siguientes videos:
Referencias
http://es.wikipedia.org/wiki/Frecuencia_de_recombinaci%C3%B3n
La variedad de Especies y su relación con Ligamiento y Entrecruzamiento
Publicado por: Kimberly Belandria

Entrecruzamiento a) y b) El entrecruzamiento se inicia cuando se aparean las cromátides homólogas, al inicio de la meiosis I. Luego se produce la ruptura de las cromátides y los extremos de cada una de ellas se unen con los de su homóloga. Asi, los alelos se intercambian entre los cromosomas. c) Como resultado de este proceso, los cromosomas homólogos tienen combinaciones de alelos diferentes de las iniciales. Dando la variabilidad de las especies.
Los cromosomas sexuales, además de los genes que determinan el sexo, tienen también otros genes que no tienen nada que ver con los caracteres sexuales. Estos genes son los genes ligados al sexo.
En la especie humana, el cromosoma Y, al ser de menor tamaño, posee menos información que el cromosoma X. Esta es la razón de que la mayoría de los caracteres ligados al sexo que se conocen sean caracteres ligados al cromosoma X. Así, en el cromosoma X se han detectado hasta 150 loci, algunos de ellos portadores de ciertas anomalías.
Herencia ligada al cromosoma Y
Un gen ligado al cromosoma Y se manifestará en todos los hombres que lo lleven y sólo en los hombres, independientemente de que sea dominante o recesivo. Entre los pocos casos que se conocen de anormalidad hereditaria ligada al cromosoma Y tenemos la hipertricosis del pabellón auricular. Se trata de un carácter que se manifiesta por la aparición de pelo en el pabellón de la oreja.

– Herencia ligada al cromosoma X
Los genes dominantes ligados al cromosoma X son muy poco frecuentes. Se trata de un tipo de herencia que se caracteriza por que los varones afectados transmiten el carácter a todas sus hijas y a ninguno de sus hijos. Las mujeres afectadas lo transmiten a la mitad de sus hijos y a la mitad de sus hijas. Un ejemplo de este tipo de herencia es la hipofosfatemia (raquitismo que no cede con la administración de vitamina D).
Los genes recesivos ligados al cromosoma X sólo se manifiestan en la mujer, en el caso de que estén en homocigosis, en el hombre se manifestarán siempre.
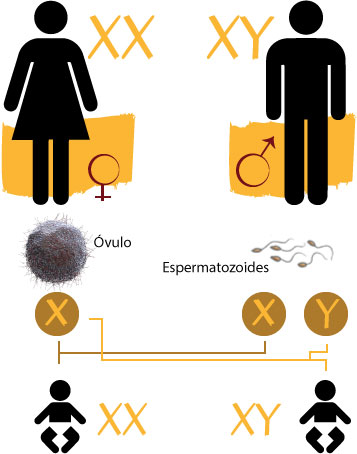
Referencia:
http://web.educastur.princast.es/proyectos/biogeo_ov/2BCH/B4_INFORMACION/T410_GENETICA
































![Drosophila_melanogaster[1]](https://i0.wp.com/upelbiogenetica.wordpress.com/wp-content/uploads/2015/01/drosophila_melanogaster1.jpg?w=391&h=261&ssl=1 "Drosophila_melanogaster[1]")


















